
论文:三个不同品系中华鳖形态差异分析
三个不同品系中华鳖形态差异分析
梁宏伟1,2,曹力欢1,2,李 翔3,童苗苗1,蒋业林4,李 忠1,罗相忠1,邹桂伟1
(1.中国水产科学研究院长江水产研究所,武汉 430223;2.华中农业大学水产学院,武汉 430223;3.安徽省喜佳农业发展有限公司,安徽蚌埠 233700;4.安徽省农业科学院水产研究所,合肥 230031)
通过测量中华鳖(Pelodiscussinensis)淮河品系(3龄)、黄河品系(8月龄)和日本品系(8月龄)的10项形态参数,对不同品系和性别的形态差异进行分析。主成分分析表明,3个品系4个主成分的累计贡献率为80.5%,其中第1主成分是中华鳖的体型因子,贡献率最大,3个品系的形态差异主要体现在体型上。通过逐步判别法建立了3个品系的判别函数,淮河、黄河和日本品系的判别准确率分别为91.7%、75.0%和73.8%,淮河鳖的形态与其他品系的中华鳖存在较大的差异。3个品系中不同性别的判别分析显示,淮河品系中华鳖雌雄判别准确率也较高,综合判别率为88.3%,判别效果最好,建立的判别函数可初步用于不同品系不同性别中华鳖的鉴定。中华鳖形态特征的差异分析为今后中华鳖种质资源鉴定和新品种(系)的选育提供了参考依据。
中华鳖(Pelodiscussinensis);形态差异;主成分分析;判别分析
中华鳖(Pelodiscussinensis)属爬行纲、龟鳖目、鳖科、鳖属,俗称甲鱼、水鱼、团鱼。在世界范围内,鳖主要分布于亚洲、非洲及北美,在亚洲主要分布于中国、俄罗斯东部、朝鲜、越南和日本[1,2]。在中国分布范围广泛,除西藏、青海及新疆外各省市、自治区均有分布,常见于长江流域和华南地区[1,3]。我国鳖种主要有中华鳖、山瑞鳖、砂鳖和斑鳖,其中山瑞鳖和斑鳖已经被列为保护动物,而养殖鳖种主要是中华鳖。由于我国幅员辽阔,气候差异大,因而不同地域之间形成了各自的中华鳖地理品系,如黄河品系、淮河品系、鄱阳湖品系、洞庭湖品系、台湾品系、崇明品系和太湖品系等,在外形和生产性能上也存在一定的差异,其中黄河品系生长速度最快,淮河鳖次之,太湖品系最慢[2,4]。近些年来,中华鳖养殖业得到较快发展,养殖产量在2015年已经达到34.2万吨[5],然而养殖企业(户)频繁引种现象极为普遍,加之缺乏科学的引种指导,造成了一些种群的混杂和优良种质特性的退化(如生长速度减慢、群体整齐度降低和畸形率增加等)[6]。
对于种质资源鉴定最为简单直观的就是外部形态特征鉴别。中华鳖黄河品系背部呈土黄色,体大裙边宽,生长和繁殖性能好;日本品系生长速度快,在浙江地区养殖效果良好;淮河品系背部呈土黄色,主要分布在安徽淮河流域两岸,以安徽蚌埠周边区域为代表,作为优良的地方种质资源越来越受到重视,目前该地方种的新品(系)种选育已经列入安徽省重大科技攻关项目[2,7,8]。目前对于不同品系之间中华鳖形态特征上的差异仅有外部形态和体色等描述,对于不同性别中华鳖的鉴定只有在出现明显异形特征时通过外部形态进行判别(早期个体的判别准确性差),或者在早期通过解剖、制作组织切片观察的方法进行鉴定。为了更加深入了解中华鳖淮河品系、黄河品系和日本品系以及雌雄性别之间的形态特征差异,本文旨在通过对3个品系的形态差异进行比较分析,为今后中华鳖新品种(系)的选育提供基础依据。
1 材料与方法
1.1 材料
本试验所采用的中华鳖淮河品系(简称淮河鳖)、黄河品系(简称黄河鳖)和日本品系(简称日本鳖)均采集自安徽省喜佳农业发展有限公司。淮河鳖为池塘养殖的3龄的中华鳖,黄河鳖和日本鳖均为温室养殖的8月龄中华鳖。雌雄性别通过外部形态特征和解剖观察性腺来确定,年龄均为生物学年龄。样本信息见表1。
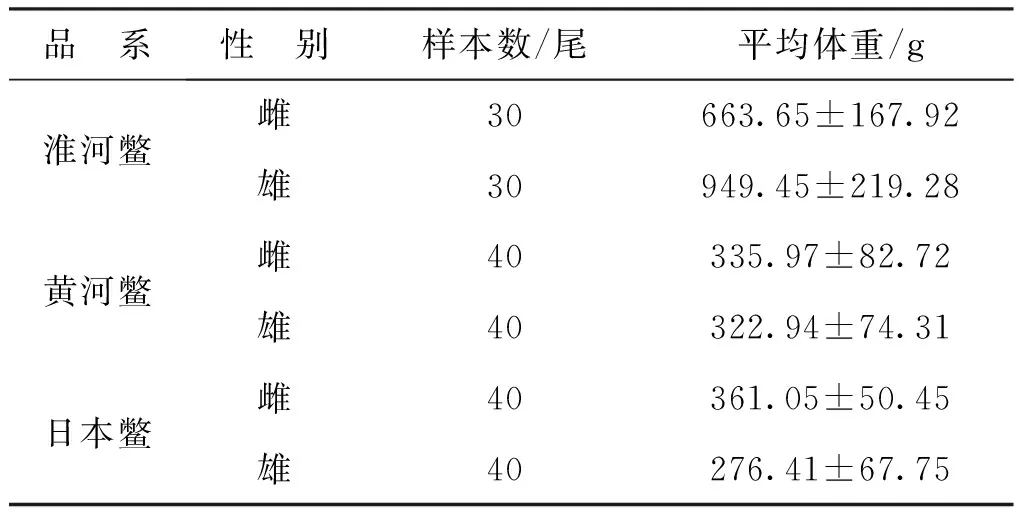
表1 3个不同品系中华鳖样本信息Tab.1 Samples information of three strains of P.sinensis
1.2 形态学测量
本研究中共对3个品系中华鳖的11项指标进行了测定,包括体重(Body weight,BW)、背甲长(Carapace length,CL)、背甲宽(Carapace width,CW)、腹甲长(Plastron length,PL)、腹甲宽(Plastron width,PW)、体高(Shell height,SH)、吻长(Snout length,SL)、吻突长(Protuberant snout length,PSL)、眼间距(Interorbital distance,ID)、吻突宽(Protuberant snout width,PSW)和后侧裙边宽(Back apron width,BAW)。体重采用电子天平进行测量(精确度为0.01g),形态学测量方法参考中华鳖国家标准(GB21044-2007),采用游标卡尺(精确度为0.1 mm)进行测量。
1.3 数据处理
1.3.1 主成分分析
由于不同规格个体对中华鳖参数值的分析会造成一定的影响,在主成分分析前,利用SPSS 22.0软件对3个品系的形态参数进行正态分布检验,然后将测得的各项参数数值分别除以对应的背甲长作为度量值予以校正,从而消除不同规格造成的影响。最后采用SPSS软件对3个品系中华鳖的形态特征数据进行主成分分析,得到各主成分特征值和贡献率及主成分载荷值。
1.3.2 判别分析
使用SPSS 22.0软件通过逐步判别法建立3个品系的判别函数和各品系雌雄群体的判别函数,据此对所有样本进行判别。判别准确率=判别正确的个体数/实测个体数×100%。
2 结果
2.1 性状参数测定
正态性检验表明,中华鳖各品系形态学数据均符合正态分布。校正后的淮河鳖、黄河鳖和日本鳖雌雄群体各性状参数列于表2,从形态参数上看,3个品系中华鳖均表现出雌性背甲宽/背甲长(CW/CL)、腹甲长/背甲长(PL/CL)和腹甲宽/背甲长(PW/CL)略大于雄性,在体型特征上存在差异。
![]()
表2 不同品系中华鳖各性状参数Tab.2 Appearance parameters of different stains of P.sinensis
2.1 主成分分析
对3个不同品系中华鳖的性状参数进行主成分分析,共获得4个主成分, 9个性状参数入选主成分的特征向量、性状特征值和累计贡献率见表3和表4。4个主成分的累计贡献率为80.5%,利用这些进行主成分分析能基本上反映所隐含的信息。第一主成分的特征根为3.93,方差贡献率为43.65%,
![]()
表3 中华鳖的主成分的特征向量Tab.3 Principal eigenvector of P.sinensis
其主要作用的是背甲宽和腹甲宽,反映了中华鳖的体型特征,可以称之为体型因子;第二主成分的特征根为1.29,方差贡献率为14.35%,其主要作用的是体高,反映了中华鳖的高度特征;第三主成分反映的是裙边的特征;第四主成分主要反映的是鳖的眼间距(方差贡献率为10.81%),称为眼部特征。淮河鳖的第一主成分的贡献率(51.47%)高于黄河鳖(35.32%)和日本鳖(33.68%)。
![]()
表4 中华鳖的特征值和累计贡献率Tab.4 Eigen and cumulative contribution rate of 3 strains of P.sinensis
2.3 判别分析
通过逐步判别分析,从9个性状比值中筛选出对中华鳖日本品系、黄河品系、淮河品系有显著贡献的6个变量:体高/背甲长(SH/CL)、腹甲长/背甲长(PL/CL)、腹甲宽/背甲长(PW/CL)、后侧裙边宽/背甲长(BAW/CL)、吻突长/背甲长(PSL/CL)和眼间距/背甲长(ID/CL),当判别函数含有这6个变量时,显著性检验结果Wilks’λ=0.571,P<0.000 1,说明利用6个变量建立的判别函数判别效果良好,具有极显著的意义。建立的判别函数如下:
![]()
表5 3个不同品系中华鳖的特征值和累计贡献率Tab.5 Eigen and cumulative contribution rate of 3 strains of P.sinensis
日本鳖=107.258 SH/CL+402.467 PL/CL+331.876 PW/CL+898.177 BAW/CL+193.996 PSL/CL+1 038.329 ID/CL-408.220;
黄河鳖=130.213 SH/CL+409.273 PL/CL+332.399 PW/CL+936.088 BAW/CL+5.955 PSL/CL+878.982 ID/CL-410.715;
淮河鳖=201.836 SH/CL+449.470 PL/CL+225.815 PW/CL+1 033.609 BAW/CL-126.280 PSL/CL+905.365 ID/CL-385.350
将中华鳖3个品系的6个形态比例的数值分别带入判别函数,得到的判别结果见表6。当判断个体中华鳖所属种群时,将所测数据校正后带入判别函数,以函数值最大的判别函数所对应的群体作为该个体所属类群。淮河鳖、黄河鳖和日本鳖的判别准确率分别为91.7%、75.0%和73.8%。淮河鳖和日本鳖之间的错判率小,在淮河鳖的判别中,没有错判为日本鳖,在日本鳖的判别中也仅有1个个体错判为淮河鳖。
为了进一步分析不同品系中华鳖雌雄个体之间的形态差异,分别对淮河鳖、黄河鳖和日本鳖的雌雄群体进行判别分析,得到腹甲长/背甲长(PL/CL)、后侧裙边宽/背甲长(BAW/CL)、吻突宽/背甲长(PSL/CL)、吻长/背甲长(SL/CL)、眼间距/背甲长(ID/CL)、体高/背甲长(SH/CL)6个显著贡献变量,从而得到判别函数为:
淮河鳖(♀)= 358.265 PL/CL+351.311 BAW/CL-9.784 PSL/CL+428.358 SL/CL+720.919 ID/CL-206.049;
![]()
表6 中华鳖3个品系的判别结果Tab.6 Discrimination among 3 strains of P.sinensis
![]()
黄河鳖(♀)= 1 152.383 PL/CL+189.629 PSL/CL+463.273 SH/CL-555.748;
![]()
日本鳖(♀)=2 084.582 PSL/CL+ 942.040 SH/CL+567.052 ID/CL-199.184;
![]()
淮河鳖和黄河鳖雌雄初始判别准确率分别与其交叉验证准确率一致(表7),淮河鳖雌性判别准确率(90.0%)高于雄性(87.6%),黄河鳖雌性判别准确率(77.5%)低于雄性(90.0%),而日本鳖雌雄性别的初始判别是判别准确率一致,均为80.0%,在交叉验证的情况下其判别准确率也基本相当。
3 讨论
主成分分析是解决多变量高维复杂系统的一种较为常见的多元统计方法,其主要是通过降维技术在尽可能少损失原有信息的前提下,将有一定相关性的多个观测指标简化为少数的几个综合因子,从而更好地反映数据特征[9,10]。该分析方法已经被广泛应用在水产动物的形态特征分析方面,并在不同群体或水系水生动物的形态特征分析方面得到良好的应用[11-13]。在中华鳖上,刘阳等[14]曾对中华鳖5个不同群体的形态学特征进行主成分分析,第一主成分反映中华鳖的整个形态变化,第二主成分反映背部、裙边和尾部的形态变化。本研究对3个中华鳖品系的9个性状比值进行主成分分析,获得4个主成分,累计贡献率达80.51%,能较好反映所测中华鳖的形态性状信息,其中第一主成分方差贡献率为43.65%,反映了中华鳖的体型特征,且包含的淮河品系信息较黄河品系和日本品系信息更为丰富,因此3个品系的形态差异主要体现在体型因子上。
![]()
表7 3个品系中华鳖不同性别判别分析结果Tab.7 Discrimination of different gender among 3 stains of P.sinensis
科研人员通常希望能够获得尽可能多的生物学数据用于科研研究,然而性状之间并非完全独立,往往彼此相关、互为影响。判别分析就是通过选择相互独立的参数来建立判别函数,不仅使判别函数的形式较为简便,而且也可以获得更好的判别效果[15]。有大量的研究表明,利用判别分析对于水生动物进行综合判别可以取得良好的结果,在不同珠江卷口鱼、小黄鱼群体、长丰鲢和选育的四倍体异育银鲫(新品种名称:长丰鲫)等方面都获得了良好的判别效果[16-19]。在中华鳖上,李思发等[20]对我国华东地区的绍兴、南京、巢湖和青岛4个群体的幼鳖和成鳖的产地进行了判别分析,结果表明对幼鳖群体的判别准确率较高(80%以上),而对于成鳖的判别准确率较差。刘阳等[14]利用9个判别贡献较大的参数建立了5个不同群体中华鳖的判别函数,判别准确率介于61.7%~89.5%之间,5个群体被分为两大支。本研究成功建立了淮河品系、黄河品系和日本品系中华鳖的判别函数,其综合判别准确率为79.1%,能够初步用于3个品系中华鳖的鉴定,特别是能有效区分中华鳖淮河品系和日本品系。三个品系中淮河鳖的判别准确率最高,也表明淮河鳖与其他两个品系的形态差异更为明显,相比较而言,黄河鳖与日本鳖的形态特征差异较小。
在许多水生动物中,雌雄个体在形态或生长上都存在一定的差异,一些种类表现出雌性生长速度快于雄性(如,半滑舌鳎和牙鲆),一些种类表现出雄性快于雌性(如,尼罗罗非鱼和黄颡鱼)。中华鳖属于雄性个体生长速度快于雌性个体的水生动物,在生产实践中雄性苗种的养殖更受欢迎,全雄或高雄性中华鳖的选育已经成为了重要的研究课题。因此,中华鳖早期性别鉴定对于中华鳖的选育具有非常重要的意义。对于中华鳖雌雄个体的鉴别,目前通常以尾的长短作为第一辨别特征,雌性个体尾短,尾端不或很少自然伸出裙边外;雄性个体尾长,能够自然伸出裙边。但是这一性别鉴定方法存在一定的不足,对于尾部特征不明显的个体或者幼鳖鉴定效果较差,特别是对于规格较小的幼鳖的判别准确性差,甚至在中华鳖苗种早期无法进行准确的性别鉴定,给科学研究带来了一定的困难。在银色鳗和金钱鱼等鱼类中成功建立了雌雄判别函数,雌雄判别率分别达到85.96%和88.5%,建立判别函数用以性别鉴定是可行的[21,22]。本研究建立了中华鳖淮河品系、黄河品系和日本品系雌雄判别函数,对于淮河品系中华鳖性别判别准确率较高,黄河品系雄性判别准确率也比较高,而日本品系中华鳖雌雄性别判别准确率相对较低。虽然建立的判别模型具有较好的判别效果,但是由于本研究中测定的生物学性状有限且采用的样本个体规格相对较大,因而在今后的科研工作中应尽可能采集更多的性状特征用于不同性别的判别分析,以期获得更好的判别效果;另外,本试验中建立的判别函数对于规格较小个体的性别判别效果还有待于进一步的验证研究。
[1]杨 萍,唐业忠,王跃招.中国鳖属的分类历史简述[J].四川动物,2011,30(01):156-159.
[2]张振东,刘夕姿,肖友红.我国中华鳖苗种产业调查与发展战略[J].渔业信息与战略,2016,31(2):98-104.
[3]王培潮.中国的龟鳖[M].上海:华东师范大学出版社,2000.
[4]蔡完其,李思发,刘至治,等.中华鳖七群体稚鳖—成鳖阶段养殖性能评估[J].水产学报,2002,26(5):433-439.
[5]农业部渔业渔政管理局.2016中国渔业统计年鉴[M].北京:中国农业出版社,2016.
[6]何 力.我国中华鳖种业的基本状况[J].渔业致富指南,2015,(20):61-62.
[7]赵春光.日本鳖生物学特性与繁养新技术(一)[J].科学养鱼,2008,(1):12-13.
[8]李思发,蔡完其,刘至治,等.中华鳖七群体体形和腹部黑斑图案的差异比较[J].水产学报,2004,28(01):15-22.
[9]王 芳.主成分分析与因子分析的异同比较及应用[J].统计教育,2003,(5):14-17.
[10]李靖华,郭耀煌.主成分分析用于多指标评价的方法研究-主成分评价[J].管理工程学报,2002,16(1):39-43.
[11]杨 慧,朱文彬,董在杰,等.3个红罗非鱼群体的形态差异分析[J].上海海洋大学学报,2015,24(5):678-684.
[12]康 伟,吴 廉,刘金生,等.图门江水系绒螯蟹的形态差异与遗传混杂[J].中国水产科学,2016,23(3):555-564.
[13]黄 斌,张斯荷,杨艳磊.华中与华南地区黄缘闭壳龟形态学差异[J].信阳师范学院学报(自然科学版),2013,26(2):240-244.
[14]刘 阳,赵 建,朱新平,等.中华鳖5个不同群体的形态差异分析[J].大连海洋大学学报,2013,28(2):174-178.
[15]李朝霞,李 健,王清印,等.中国对虾“黄海1号”选育群体与野生群体的形态特征比较[J].中国水产科学,2006,13(3):384-388.
[16]赵 建,朱新平,陈永乐,等.珠江卷口鱼不同地理种群的形态变异[J].动物学报,2007,53(5):921-927.
[17]孙 琛,王肇霖,刘 峰,等.小黄鱼4个地理群体形态差异分析[J].浙江海洋学院学报(自然科学版),2016,35(3):186-194.
[18]梁宏伟,李 忠,罗相忠,等.长丰鲢与长江鲢形态差异与判别分析[J].水生生物学报,2015,39(5):1059-1064.
[19] Li Z,Liang H W,Wang Z W,et al.A novel allotetraploid carp strain with maternal body type ang growth superiority [J]. Aquaculture,2016,485: 55-63.
[20]李思发,李晨虹,吕国庆,等.华东地区中华鳖地方群体形态差异初步研究[J].中国水产科学,1997,(S1):2-8.
[21]郭弘艺,魏 凯,谢正丽,等.长江口银色鳗的形态指标体系及其雌雄鉴别[J].水产学报,2011,35(1):1-8.
[22]吴 波,张敏智,邓思平,等.金钱鱼雌雄个体的形态学差异分析[J].上海海洋大学学报,2014,23(1):64-69.
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!
