
论文:褶皱臂尾轮虫土池大规模培育中酵母维生素替代鸡粪的应用效果

褶皱臂尾轮虫土池大规模培育中酵母维生素替代鸡粪的应用效果
朱天,徐世陈,蔡萌,刘洋,韩萃,陈德芝,杨家新*
(1.南京师范大学海洋科学与工程学院,江苏 南京 210023;2.射阳诚信长江蟹生态育苗场,江苏 盐城 224000)
褶皱臂尾轮虫因其体型小,游动缓慢,且能在短时间内进行快速培育,使其成为许多海产经济动物的开口饵料。自2001年以来,在我国沿海地区,利用鸡粪为基肥并结合其他外源物和鸡粪追肥培养饵料轮虫的土池生态育苗模式,已经成为我国河蟹育苗饵料生产的主推模式。轮虫主要以微藻为食,在底泥中的轮虫休眠卵受温度诱导催发后,在合适的微藻密度下,轮虫的种群密度能够在较短的时间内快速增长。鸡粪因其富含大量氮、磷等生物基本元素,能够有效地提高池塘的初级生产力,促进微藻的生长。因此,鸡粪常常作为轮虫大规模培育中培育池的主要基肥。然而,由鸡粪介导的氮磷输入易导致水体的富营养化,且在高浓度的氮磷胁迫下,轮虫的生长与繁殖也会受到负面影响,从而影响轮虫的产量。因此,需要一种更为清洁、高效的施肥/追肥方式来优化土池培育轮虫。
通过培养微藻喂养轮虫的方式,很难在轮虫大规模培育中支撑起轮虫的摄食需求,因此需要与其他营养物相结合来实现轮虫的规模化培育。面包酵母富含蛋白质、氨基酸等营养物质,是一种广泛使用且极具潜力的轮虫饲料。酵母的含氮量较低,将酵母进行活化后可用于轮虫的连续培养,且不易造成水体污染。因此,酵母可以作为轮虫的补充饲料用于投喂,减少培育池内用于培养微藻的鸡粪投入量。
此外,在轮虫大规模生产中,轮虫达到高峰期后,时常出现“种群崩溃”现象,严重影响了轮虫产量与稳定性。针对这一现象,一些学者认为添加适量的维生素有利于防止轮虫的种群崩溃,提高轮虫的产量。目前常用于轮虫培育的维生素主要有维生素E(V)、维生素C(V)、维生素B12(V)等,维生素与酵母混合投喂能够有效地使其在轮虫体内富集。现参考国内外文献,选用V、V和V3 种维生素,制定合适的添加量,与酵母制成新型投喂品,对轮虫培育池追肥,并与使用鸡粪追肥的培育方式进行比较,探究酵母维生素在轮虫培育池实际生产中的应用效果。
1 材料与方法
1.1 池塘
试验地位于江苏省盐城市射阳诚信长江蟹生态育苗场,选择2口土塘(PA、PB)作为轮虫培育池。面积均为7m×35 m,平均深度均为1.5m,底质均为黏性土壤。分别在2口池塘的上风口、下风口和池塘中央设置3个采样点。PA塘、PB塘的水源均为经过过滤处理的天然海水。使用前对2口池塘清塘消毒,泼洒质量浓度为30~50g/m的漂白粉,暴晒2~3d后投入使用。其中PA塘使用维生素和酵母混合物进行追肥,PB 塘使用发酵鸡粪追肥。
1.2 酵母维生素混合物
PA 塘、PB 塘均以鸡粪为基肥,并人工接种小球藻。PA 塘培育过程中,通过观察其浮游动物和微藻的种群密度变化,使用酵母维生素混合物追肥1 次。酵母维生素混合物配方:酵母(60 kg/hm,安琪酵母股份有限公司)和水、酵母、红糖(100∶10∶1)搅拌15 min 后,将其置于阴凉处活化30 min,取上清液与维生素混合。维生素添加量为:V100 g(郑州中牧饲料有限公司公司生产)、V200 g(强胜饲料有限公司生产)、V200 g(强胜饲料有限公司生产)。PB 塘同日使用发酵鸡粪追肥1 次,用量为60 m/hm。
1.3 样品采集
采样时间固定为每日08:00,使用13 号、25 号浮游生物网采集轮虫和浮游植物进行定性定量分析。采集10 L 水样,过滤浓缩至50 mL,制成混合水样。水温(WT)、溶解氧(DO)、pH 值、电导率(Cond)、盐度(SAL)、叶绿素a(Chl-a)采用多参数YSI 水质分析仪测定和希玛pH 计进行现场测定。使用HACH DR 900 分析仪配合DBR 200 消解器测定水体的铵态氮(NH-N)和总磷(TP)。
1.4 数据处理与分析方法
使用SPSS 21.0、R 软件进行数据检验和池塘环境因子的主成分分析(PCA)。采用单因素方差分析(ANOVA),对轮虫生命表参数和种群增长参数进行正态分布检验和方差齐性检验,若数据符合正态分布和方差齐性,则对数据进行单因素方差分析,组间多重比较采用Turkey 检验,对于不符合方差齐性的数据,使用Tamhane’s T2 检验进行多重比较。使用Origin 2019b 绘图。
2 结果与分析
2.1 浮游生物
检测轮虫培育池的浮游植物种类,有利于分析培育池内轮虫饵料的组成以及水环境。轮虫高峰期内,培育池内共检测出藻类5 门9 种(表1)。轮虫培育前期(1~4 d),池塘的藻类组成主要以绿藻(小球藻)、硅藻(中华盒形藻、格式圆筛藻)和黄藻(海绿球藻)为主,其中小球藻为人工接种的藻种;培育后期(7~10 d),2 口池塘除小球藻外,裸藻(尖尾裸藻、静裸藻)出现的频次较高。

表1 培育池内浮游植物种类和出现频次①

续表
2.2 轮虫密度与携卵率
轮虫种群密度和携卵率变化见图1(a)(b)。PA 塘的轮虫初始密度(55.25±2.82)ind./mL 与PB塘(55.00±2.47)ind./mL 相近。试验第2 天,对2口池塘进行追肥,PA 塘轮虫密度在追肥1 d 后(第3天)明显增长,轮虫密度达到峰值(66.5±4.18)ind./mL,此后密度有所下降,在培育期的3~10 d,PA 塘轮虫密度均显著高于PB 塘(P<0.05)[图1(a)]。轮虫的携卵率代表了轮虫种群的繁殖能力,由图1(b)可见,试验1~2 d,2 口池塘内轮虫携卵率并无显著差异,而当使用不同肥料进行追肥后,PA 塘的轮虫携卵率显著高于PB 塘(P<0.05),此后5~7 d PA 塘的轮虫携卵率显著高于PB 塘(P<0.05),8~10 d 2 口塘轮虫携卵率无显著差异(P>0.05)。
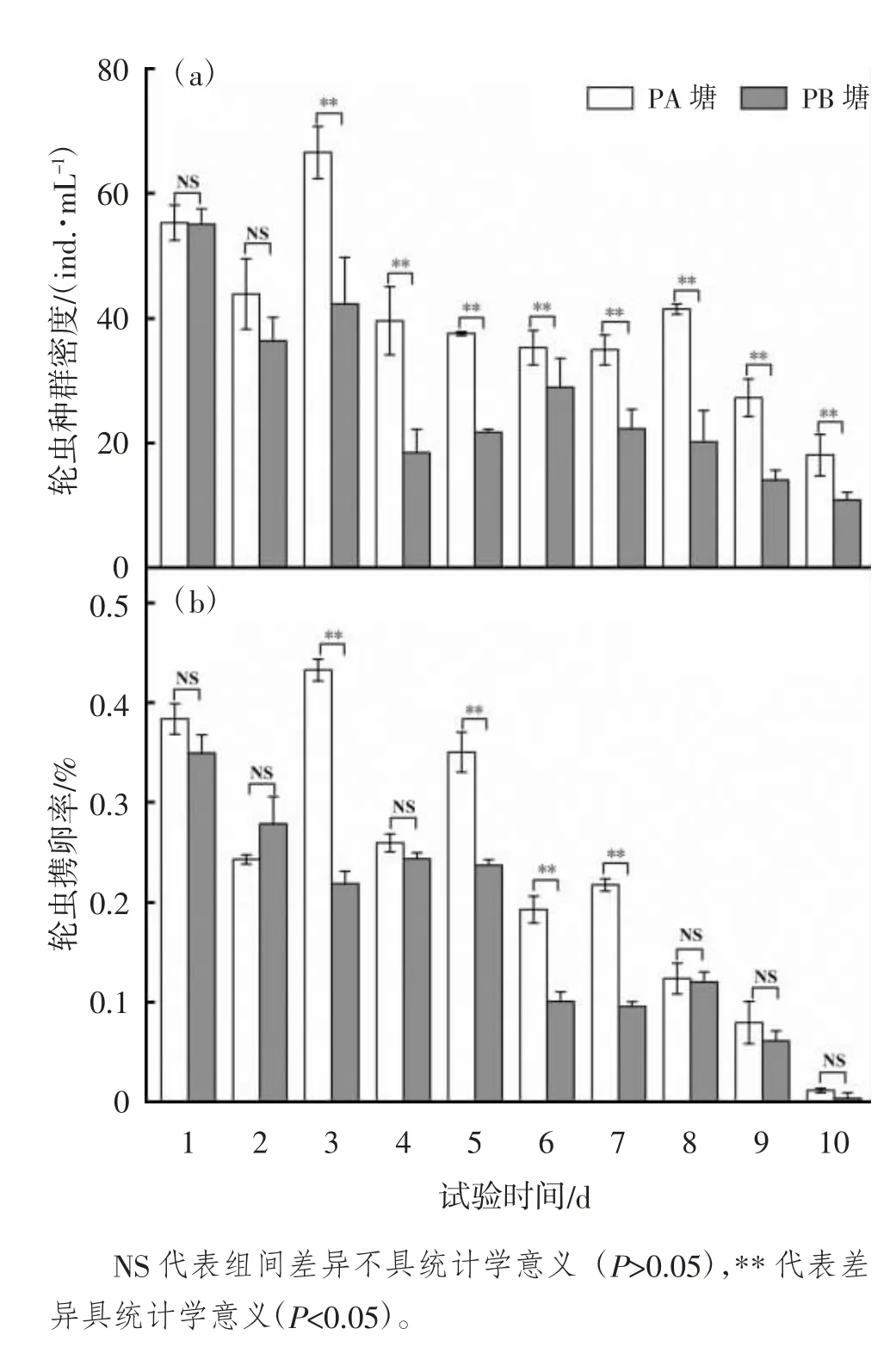
图1 轮虫种群密度和携卵率变化
2.3 环境因子主成分分析
试验期间对PA 塘和PB 塘理化因子每日检测,其变化范围见表2。使用主成分分析来实现对两池环境因子的时空变化的可视化。用于解释主成分的前2 个变量(PC1,PC2)用于绘制二维散点图,其结果见图2(a)(b)。PA 塘池塘环境因子[图2(a)] 的前2 个主成分解释了总变量的90.88%,PC1 解释了83.27%,PC2 解释了7.61%。培育池1~3 d,4~6 d,7~10 d 这3 个阶段拥有不同的水环境主成分。其中DO 和NH-N 在PC1 上具有较高的特征值,分别为0.96 和-0.15;Cond、SAL 和NH-N在PC2 上具有较高的绝对特征值,其特征值分别为0.88,0.33 和-0.31。PB 池池塘环境因子[图2(b)]的前2 个主成分解释了总变量的88.25%,PC1 解释了59.27%, PC2 解释了28.98%。根据图2(b),可以将培育池的水环境分为1~6 d 和7~9 d 2 个阶段,NH-N、DO 和SAL 在PC1 上拥有较高的特征值,分别为-0.73,0.61 和-0.24;NH-N、DO 和Cond 在PC2 上具有较高的特征值,分别为-0.69,-0.67 和0.24。
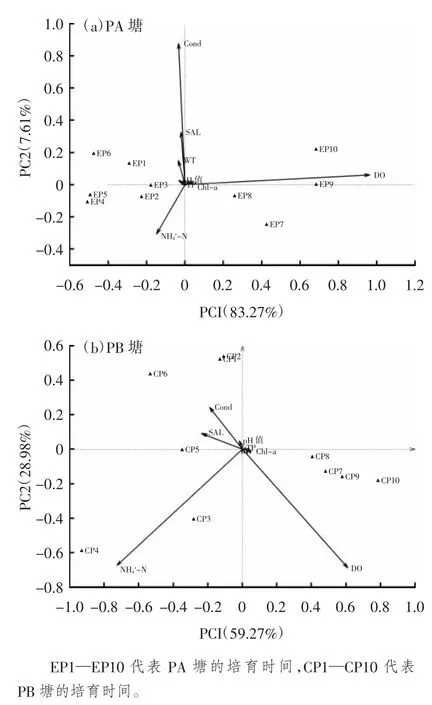
图2 环境因子主成分分析
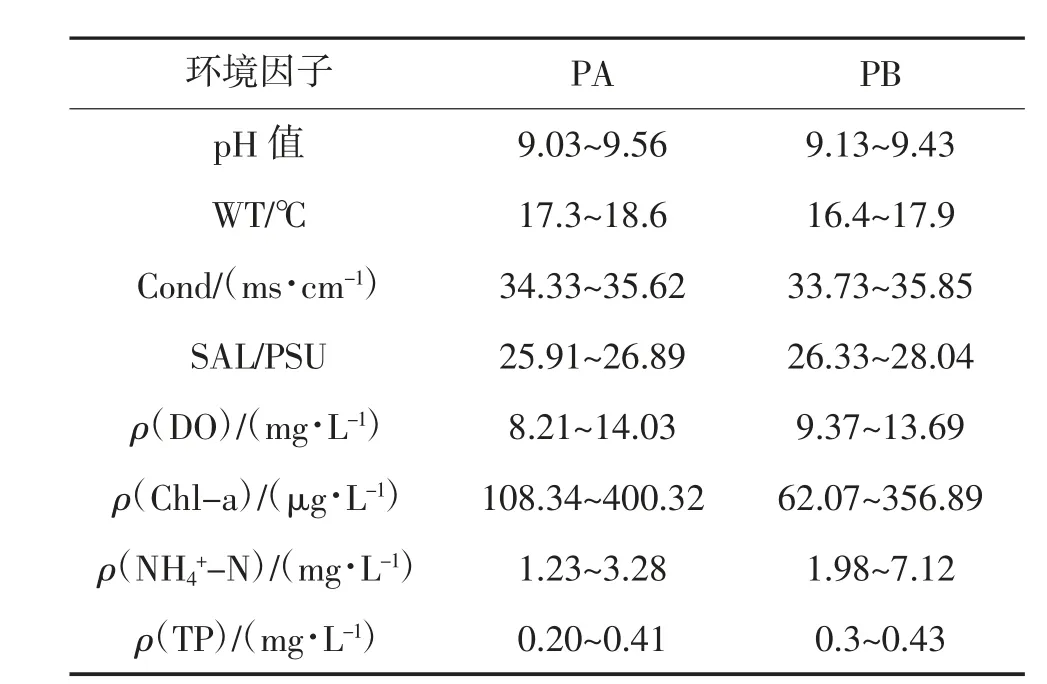
表2 2 口池塘水质环境因子的变化范围
2.4 轮虫密度与环境因子相关性分析
通过对轮虫密度与池塘环境因子之间的相关性分析发现,影响PA 塘轮虫密度的主要环境因子为pH值、Chl-a 和NH-N,Pearson 相关性系数分别为0.64、-0.72和0.83;影响PB 塘轮虫密度的主要因子为DO 和TP,其Pearson 相关性系数分别为-0.63 和0.76(表3)。
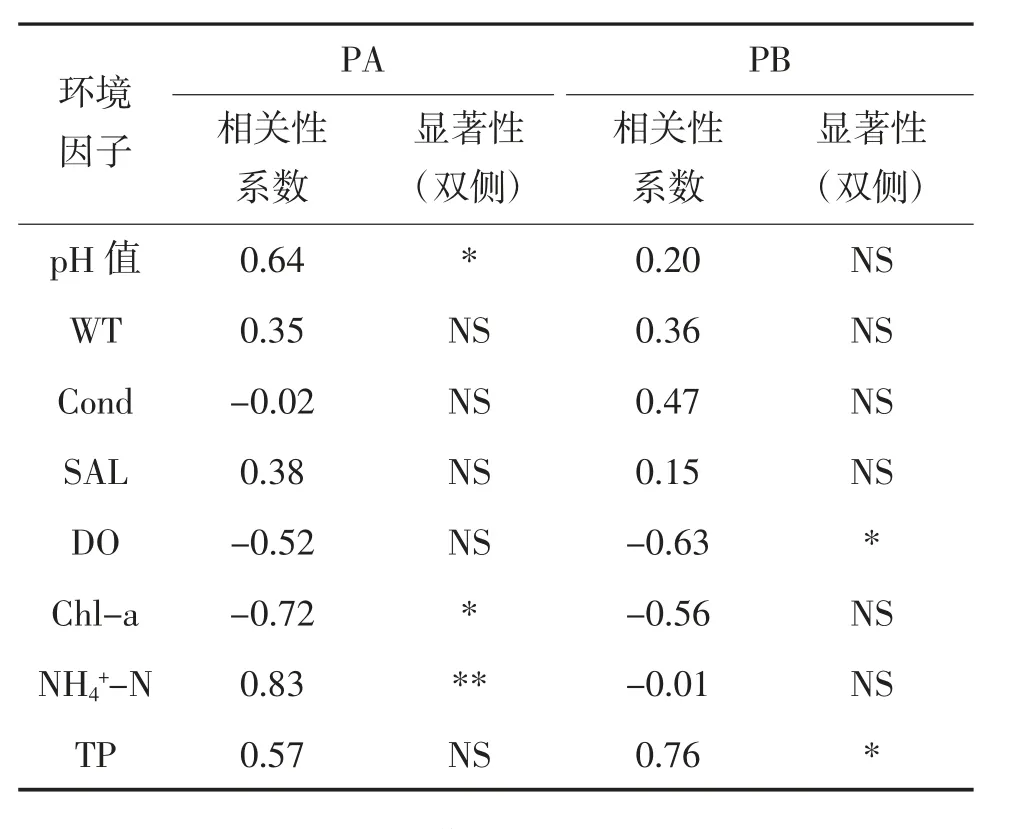
表3 轮虫密度与环境因子间相关性分析①
3 讨论
随着中华绒螯蟹集约化育苗产业的扩大,溞状幼体短期内对轮虫的集中需求,导致了对生物饵料轮虫的大量需求。大规模培育轮虫的过程中,轮虫的种群密度受到食物浓度、环境因子以及营养水平等多种因素的协同影响。不同的饵料密度会导致不同的轮虫种群增长;此外,不同水平的盐度、WT、DO、NH-N 和TP 也会影响轮虫的种群密度。因此,2 种投喂方式对土池轮虫培育的影响,应当从多个方面考虑。
鸡粪的汇入,能够为池塘提供大量的氮磷等基本元素,增加水体的初级生产力,从而促进藻类的生长,以供轮虫摄食。微藻作为轮虫的基础饵料,是天然水体中轮虫的主要食物与营养来源。区别于鸡粪施肥促进藻类生长以喂养轮虫的间接培育方式,酵母可被轮虫摄食,且含氮量极低,在轮虫的连续培养中,不易造成水体污染,这2 种施肥方式,也会导致不同的水环境与浮游植物组成结构改变。研究表明,轮虫具有摄食选择性,在绿藻、硅藻和裸藻混合的水环境中,轮虫主要以摄食绿藻(小球藻,微绿球藻等)为主。其中,海水小球藻富含轮虫所需的各类营养以及丰富的高价不饱和脂肪酸(HUFAs),能够强化轮虫的营养价值,被认为是最适合用于培养轮虫的藻类之一。因此,在土池培育中,小球藻常以人工接种的方式用于投喂轮虫。该研究的结果中,PB 塘较PA 塘的浮游植物组成更为丰富,而多种藻类共存的水环境不利于单一藻种的生长。因此,使用酵母与维生素进行追肥相比于鸡粪追肥,在培育池浮游植物的组成结构上更加有利于培育轮虫。
小球藻与酵母是培育轮虫的2 种常见饲料,小球藻用于投喂轮虫,能够为轮虫提供较为全面的营养,而与小球藻相比,酵母缺乏幼苗生长所需的HUFAs,如二十碳五烯酸(EPA)、二十二碳六烯酸(DHA)等。因此,使用酵母培育轮虫时,通常需要对轮虫进行营养强化以满足幼苗的摄食需求。文献[9,14-16]研究表明,在轮虫培育过程中添加V、V、V,能够提高轮虫的抗氧化能力,并促进轮虫种群的增长与繁殖力。Tako研究表明,同时添加维生素和小球藻,能够有效防止HUFAs 的氧化,增加轮虫HUFAs 的含量。因此,基于这些学者的研究,选用酵母与维生素进行混合投喂,以求达到更好的轮虫培育效果。本研究中,轮虫种群密度的差异始于追肥日(3 d),PA 塘中轮虫的种群密度于3~10 d 显著高于PB 塘(P<0.05);轮虫携卵率的显著差异出现在追肥后的3~6 d,PA塘中轮虫的携卵量显著高于PB 塘(P<0.05),这与预期的结果相一致,PA 塘的轮虫在产量与繁殖能力上皆显著高于PB,有效实现了轮虫增产。
现使用主成分分析(PCA)和Pearson 相关性分析的方法,分析培育池的水环境,揭示2 口池塘中的环境因子变化,以及环境因子与轮虫产量之间的关系。由PCA 的结果可以得出,2 口池塘1~6 d 的水环境主成分均与NH-N 含量有较高的相关度,而7~10 d 的主成分受DO 的影响较大。PA 塘中轮虫密度与环境因子间的相关性分析表明(表3),培育期间,PA 塘内的轮虫密度与NH-N 含量呈显著正相关(P<0.05);而PB 塘中NH-N 含量与轮虫密度的相关性并不显著(P>0.05),且相关性系数为负值(-0.01),TP 与DO 更多地介导了轮虫密度的变化。蔺丽丽等认为,水体中的NH-N 含量超过5 mg/L,便会对壶状臂尾轮虫(Brachionus urceolaris)产生毒害作用,导致轮虫的种群增长与繁殖力降低。本试验中,PB 塘中NH-N 的峰值达到了7.12 mg/L(表2),超过了5 mg/L,相关性分析的结果也能够印证这一观点。因此,在培育期间,PB 塘水环境中的NH-N 含量对轮虫种群的增长产生了负面影响,这也是导致PB 塘轮虫种群密度低于PA 塘的原因之一。综上所述,在以鸡粪为基肥与追肥的轮虫土池大规模培育中,使用酵母维生素混合物追肥代替鸡粪追肥的方式,能够使培育池拥有更好的水环境。该方法能够减少一半鸡粪的投入量,从源头减少氮磷的汇入。此外,从生物防治污染的角度来说,更高的轮虫产量,也代表能够以生物的方式实现氮磷等基本营养元素的固定,并从水体中移出,防止更多的营养物流入天然水域,有效缓解轮虫培育池带来的尾水污染。
4 结语
褶皱臂尾轮虫(B.plicatilis)大规模土池粗放式培育中,使用酵母维生素混合物追肥与直接使用鸡粪追肥的方式相比,酵母维生素混合物方式有利于培育池内饵料微藻的培养,提高轮虫密度与繁殖能力,增加了轮虫产量。此方法减少了一半的鸡粪投入量,能够在轮虫培育池内维持更好的水环境,有效降低轮虫培育带来的污染,操作简易,适合在沿海生物饵料生产企业中进行推广。
声明:本站未注明出处的转载文章是出于传递更多信息之目的。若有未注明出处或标注错误或侵犯了您的合法权益,请与本网站联系,我们将及时更正、删除,谢谢!
欢迎关注本站(可搜索)"养鱼第一线"微信公众帐号和微信视频号"养鱼第一线刘文俊视频号"以及头条号"水花鱼@渔人刘文俊",将会定期向你推送信息!
