
渔药药物效应动力学——药物-受体学说
1.占领学说 C1ark于1926年提出的占领学说认为:药物效果与药物占领受体数目成正比,药物占领受体数目取决于细胞表面受体的密度和受体周围的药物浓度,当全部受体被占领,即达到Emax。占领学说只适用于激动药,但不能解释2个激动药激动同样受体而产生不同的Emax。
1954年Ariens修正了占领学说,他把决定药物与受体结合时产生效应的能力称为内在活性。药物与受体结合不仅需要亲和力,而且还需要内在活性才能激动受体而产生效应。只有亲和力而没有内在活性的药物,虽可与受体结合,但不能激动受体故不产生效应。
1956年Stephenson又提出,药物只占领小部分受体即可产生最大效应,未经占领的受体称为储备受体。因此,当不可逆性结合或其它原因而丧失一部分受体时,并不会立即影响最大效应。进一步研究发现,内在活性不同的同类药物产生同等强度效应时,所占领受体的数目并不相等。激动药占领的受体必须达到一定阈值后才开始出现效应。当达到阈值后被占领的受体数目增多时,激动效应随之增强。阈值以下被占领的受体称为沉默受体。
2.速率学说 1961年Paton 提出了受体速率学说。该学说认为影响药物效应的大小的主要因素不是受体被占领数量的多少,而是药物分子与受体的结合速率。认为每当一个药物分子和受体相碰撞时即可产生一定量的刺激,并能被传到相应效应器而产生效应。使用这一学说的简单之处在于,仅以受体-药物结合和解离速率参数即可推算结果,无须使用内在活性和效能等参数。但问题在于这一学说无法解释药物与受体多种类型的相互作用。故这一学说被使用的范围并不大。
3.二态模型学说 Kar1in A.和Changenx JP.分别提出了药物受体作用的两态学说,又称变构学说。它们都认为受体存在活性状态(R*)和非活性状态(R)。两者均可与药物结合,而且活性和非活性受体之间也可以相互转化,二者处于动态平衡。激动剂主要与R*结合形成R*A,而拮抗剂则主要形成RB。实际上激动剂和拮抗剂分别推动R*和R的转换向着各自方向移动。这两种状态的受体之比例及两种状态受体与药物的亲和力大小决定着生物效应的大小。这种模型考虑到了受体与药物间相互作用导致的受体活性改变,更接近于实际的药物-受体反应状况,但用以直接说明药物与效应之间的定量关系尚不完备。
4.诱导契合学说 Kosh1and等提出的诱导契合学说,可以较好地解释配基与受体结合的实际过程。即药物与受体蛋白结合时,可诱使受体的空间构象发生可逆改变,而且这种结构变化即可导致生物效应。而且这种学说也可以解释药物与受体之间的协作效应。
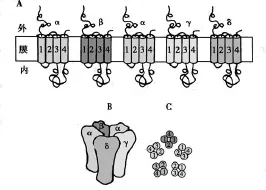
图 N2型乙酰胆碱受体阳离子通道分子结构示意图
需要注意的是,上述这些受体学说大多只能在某种特定的条件或范围内解释受体与药物的作用和受体与效应之间的关系,均存在一定的局限性。
声明:本站属公益性没有商业目的的网站,上列文章仅供个人学习参考。本站所发布文章为原创的均标注作者或来源,未经授权不得转载,许可转载的请注明出处。本站所载文章除原创外均来源于网络,如有未注明出处或标注错误或侵犯了您的合法权益,请及时联系我们!
欢迎关注本站(可搜索)"养鱼E线"微信公众帐号和微信视频号"养鱼一线"以及头条号"水花鱼@渔人刘文俊"!
